
Using the immune system optimally
Poultry diseases contribute to an important set of problems within broiler production systems. The short cycles in broiler production realised through improved genetics and diet have led to a limited recovery time after disease outbreaks. It’s the broilers immune system which has tremendous potential.
Diseases and the short recovery time a diseased broiler has in its relatively short lifetime, has consequences that include animal welfare issues, performance and economical losses. On top of that there can be a negative impact on meat safety, human health, and the concern of antibiotic use and resistance. Undoubtedly, disease outbreaks need to be prevented and not cured. Prevention and management strategies are of great importance. Biosecurity is a well-known concept, and described as a programme designed to prevent the exposure of birds to diseases by reducing the introduction and spread of pathogens into and between the farms. Vaccination programmes can be taken as an example. Despite the extensive biosecurity measures taken, the threat of pathogens will always persist.
Therefore, there is a constant drive to use specific, clearly defined ingredients to help producers reach both aims of reducing antimicrobials while maintaining animal health and productivity. An overlooked potential to help the situation is the animal’s own immune system, as people tend to forget the immune system is a magic doctor!
Immune modulation
An area, not systematically targeted yet in disease prevention, is the immune system of an animal. By priming the immune response with a feed additive, it is possible to enhance the natural defence system at a minimal energy cost and consequently increasing resistance to diseases. And even without a disease challenge there might still be a need to assist the immune system. Several well-known diseases have a down-regulation of the immune system as part of their strategy of infecting a host. Even some of the strategies of a biosecurity programme, come at a cost in terms of productivity, such as intensive vaccination programmes. Not to forget young animals, whose immune system is immature, but nonetheless present. By supporting the immune system during stressful events, there is a potential to lessen all those negative effects on the immune system. Of course, modulating the immune system always comes with the fear of serious energy costs and loss in feed conversion.
Figure 2 – Beta-(1,3)-glucan is inducing a dose responsive and consequently predictable release of immune modulating cytokines (Interleukin-6, -8, and -18). A, B, C, and D is equivalent to 0, 50, 200 and 2000 g per ton of feed respectively.

The key is to target those species or ages that genuinely need support and to standardise the active ingredients and dosage to such an extent that an overregulation can be avoided. A perfect candidate for such an in-feed supplementation is a Beta-(1,3)-Glucan derived from a new and unique source; an alga, called Euglena gracilis. Euglena gracilis is a fresh water protist and is synthesising Beta-(1,3)-Glucan as its storage reservoir of carbon. This storage molecule is highly bioavailable as it is located in the cytoplasm of the alga and it compromises of a chain of glucose molecules connected by beta-glycosidic bonds between 1 and 3 carbons. In animals, these typical 1,3 linked glucose molecules are recognised by a receptor (Dectin-1) located on immune cells, especially on antigen presenting cells like macrophages and induce a further immune modulating effect.
How to improve the immune system
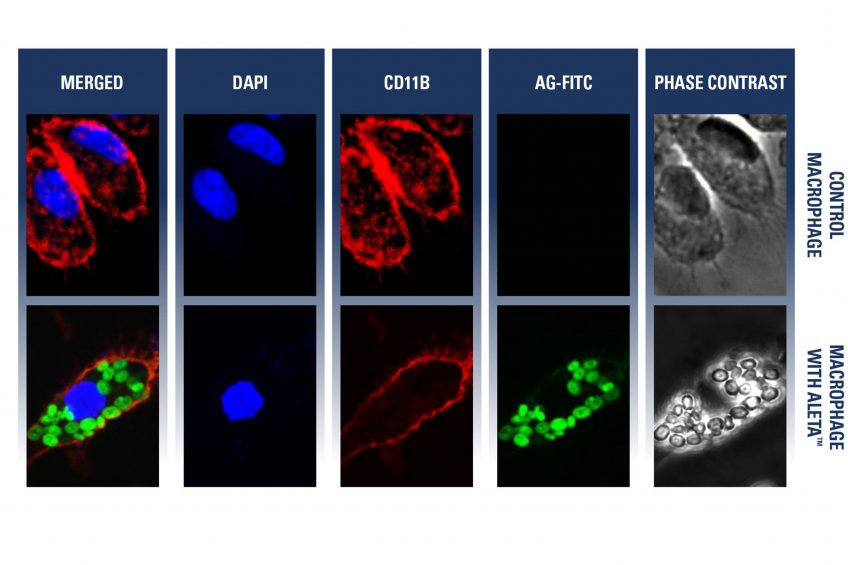
An ideal immune modulating substance needs to help induce an immune response to pathogens entering the animal’s body. Therefore, the immune response cascade needs to be respected and the following characteristics are required: the candidate ingredient must be recognised by antigen presenting cells (macrophages/dendritic cells) and induce a secretion of cytokines (responsible for intercell communication), resulting in recruitment and activation of other immune cells combatting the pathogen. In this article, trial data are summarised showing the potential immune modulating capabilities of an algal derived beta-(1,3)-glucan (Aleta). During a first experiment, mouse macrophages were collected from mouse intestine and were incubated with fluorescein isothiocyanate (FITC)-linked beta-(1,3)-glucan preparations for 2 hours and stained for macrophage marker (CD11b) and nucleus marker (DAPI) and imaged.
Secondly, mice were given FITC-linked beta-(1,3)-glucan, equivalent to 50, 200 and 2000 g per ton of feed respectively, for seven consecutive days. On day eight, macrophages were isolated from small intestine, cultured, and the supernatants were tested for the expression of various cytokines: IL-6, IL-10 and IL-18. During those experiments it was demonstrated that these beta-(1,3)-glucan is recognised and phagocytosed by antigen presenting cells (macrophages) (Figure 3) and is consequently inducing a release of immune modulating cytokines like IL-6, a pro- and anti-inflammatory cytokine, IL-10, an anti-inflammatory cytokine, and IL-18, a pro-inflammatory cytokine (Figure 2). The algal derived beta-glucan triggers a balanced immune response, as both pro-inflammatory and anti-inflammatory cytokines are secreted: activation but no overstimulation of the immune system. The immune modulating effect was proven!
Figure 3 – A significant increased APC frequency and MHC II expression in all beta-(1,3)-glucan supplemented groups compared to non-supplemented control groups.

Broiler trial
The data needed to be confirmed in livestock, and consequently a broiler trial was performed. 320 one-day old COBB 500 male broilers were divided over eight cages per treatment. The experiment consisted of four treatments: a non-infected, non-supplemented group (1), an infected, non-supplemented group (2), and two infected, supplemented groups (3, 4). Aleta was added at 100 and 150 g/ton, in groups 3 and 4 respectively, to a standard starter diet fed for 28 days. On day 14, all birds except those in group 1, were orally challenged with a coccidial inoculum consisting of mixed cultures of Eimeria species: E. acervulina, E. maxima, and E. tenella. On day 20, birds from each cage were sacrificed and lesion scored. On day 28, sections of the ileum were taken for analysis of immune cell density, expressed as total antigen presenting cell (APC) frequency, and activity, measured by MHC-II (Major Histocompatibility Complex) marker staining.
The results show an enhanced recruitment and activation of antigen presenting cells in the ileum by beta-glucan supplementation, increasing the animal’s resistance to disease. This explains the significant reduction in intestinal lesions in response to the Eimeria challenge in the beta-(1,3)-glucan supplemented groups (Table 1).
Conclusion
Research shows that specific ingredients, namely beta-(1,3)-glucans derived from algae, are effectively serving as immune modulating molecules enhancing the natural defence mechanisms of the animal against diseases and consequently contributing to an improved animal health and performance. This leads to the conclusion that the broiler’s immune system is really a magic doctor and needs to be used systemically to prevent disease outbreaks, and can be an additional tool to reduce antibiotics and support biosecurity programmes.
Join 31,000+ subscribers
Subscribe to our newsletter to stay updated about all the need-to-know content in the poultry sector, three times a week.







 Beheer
Beheer WP Admin
WP Admin  Bewerk bericht
Bewerk bericht